522427198603050819
(威宁县毕绿生态绿色产业发展有限公司 553100)
摘要:森林生态系统林分结构和生物多样性对多种生态功能至关重要,然而,林分结构和生物多样性如何驱动生态系统服务演变仍不清晰。为阐明它们之间的作用关系,研究以石漠化区阔叶混交林(MBF)、阔叶纯林(BF)、针阔混交林(MCBF)、针叶林(CF)为研究对象,通过野外监测和室内测定了4种森林治理模式的空间结构、生物多样性与服务特征,利用冗余分析探讨了空间结构(角尺度(Wi)、混交度(IcI)、竞争指数(Mi))和生物多样性(物种多样性、功能多样性)对供给、支持和调节3种生态系统服务的影响。结果表明:与其他3种治理模式相比,MCBF的竞争强度(48%~304%)和混交度更高。在物种多样性方面,MCBF的丰富度指数和均匀度指数分别是CF和MBF的2.07和1.07倍、1.95和1.3倍。功能多样性中,MCBF的Rao指数分别是CF、MBF和BF的1.25、1.02和1.4倍。在生态系统服务方面,MCBF土壤碳氮含量、水源涵养、碳储存和木材产量均显著高于其他治理模式。冗余分析显示竞争指数和混交度与水源涵养、固碳释氧和木材产量呈显著正相关,而物种和功能多样性又与土壤碳氮含量、水源涵养、碳储存和木材产量显著相关。研究表明高混交度和竞争强度会加剧物种生态位分化,有助于提升森林群落物种多样性和功能多样性,进而促进调节服务的提升。总之,森林空间结构通过驱动生物多样性作用生态系统服务。研究可为森林可持续经营提供理论依据。
关键词:空间结构;物种多样性;功能多样性;喀斯特石漠化
引言
喀斯特生态系统具有土层浅薄、土体破碎、水土易于流(漏)失、植被不易定植等生态脆弱特性特征,加上西南地区持续的人为干扰和资源的不合理利用,导致喀斯特植被破坏严重,生态系统退化和石漠化等问题较为突出(郭柯等,2011;Ni et al.,2015; Tong et al,m2018; 熊康宁等,2012;2016)。自然和人为引起的森林面积缩小,造成栖息地破坏,加剧了植物和动物物种多样性的丧失。使森林生态系统的结构失衡与功能退化,最终影响到森林生态系统为人类提供服务的能力。物种组成是群落最基本的特征,是形成群落结构的基础。物种多样性是生物多样性在物种水平上的表现形式,可反映生物群落结构的复杂性,可作为判断生物群落结构变化或生态系统稳定性的指标(Ghaley and Porter,2014; 孟莹莹等,2015)。因此,增加生物多样性有助于维持群落结构的稳定性,提高喀斯特森林生态系统水量调节、气候调节、水土保持等多种生态系统服务供给能力。不同的物种在生态系统中占据的生态位不同,这就导致了不同物种之间的生态功能差异。例如,高叶面积有助于减少土壤侵蚀(Coelho et al.,2018),同时对空气质量调节和土壤水分也具有正向的作用(Speak et al.,2020; Vasquez-Valderrama et al.,2020)。高的根系氮含量和根系直径也能提高固碳能力和土壤保持服务,而比根长增加会降低固碳服务和土壤保持服务(Demenois et al.,2018;Chen et al.,2019;Hao et al.,2020)。然而,喀斯特地区森林生态系统结构与生物多样性之间的相互作用关系如何对生态系统服务产生影响尚不清晰。因此厘清群落结构-功能-服务之间的关系,有助于完善石漠化森林生态系统的功能与提升服务供给能力。基于此,本文拟探讨:1)石漠化治理森林生态系统结构与生态系统服务存在什么样的关系;2)石漠化治理森林生物多样性与生态系统服务存在什么样的关系;3)森林空间结构与生物多样性如何驱动生态系统服务变化。对森林生态系统的管理与生态脆弱区生态修复提供参考,为区域可持续发展提供借鉴。
材料与方法
研究区概况
研究区位于贵州省毕节市威宁县么站镇,地处东经104°08′—104°21′,北纬26°36′45″—26°51′,面积196.7平方公里,其中耕地127367亩,林地109648亩;牧草地22950亩,森林覆盖率56.67%。平均海拔平均海拔2115米,年平均气温16.5℃左右,年均降水量700 mm左右,主要集中在6-8月,夏秋温暖湿润,春冬寒冷干燥。区内多峰丛洼地地貌,地形破碎,是典型的喀斯特高原山地生态环境。地带性植被以阔叶林、针叶林以及灌丛为主,人为扰动长期存在,导致植被存在不同程度退化。主要物种有华山松云南松、柳杉、杜鹃和火棘等。
样地选择
研究区内的森林按成因主要分为封育后的天然林和石漠化治理的人工林两种,天然林以混交林为主,人工林则以纯林为主。根据不同的群落物种组成,将森林划分为阔叶混交林(MBF)、阔叶纯林(BF)、针阔混交林(MCBF)、针叶林(CF)4种林分类型,分别探讨4种林型的空间结构特征、生物多样性特征和生态系统服务特征。每种林型选取3个(共12个)20 m×20 m的矩形样地,然后分别在每个样地的4角各划出1个5 m×5 m的矩形灌木样方和1 m×1 m的草本样方,记录样地经纬度、坡度坡向、海拔等地形因子。胸径大于2 cm的木本植物为划为乔木,小于2 cm的为灌木。
森林空间结构
于2023年7月在研究区开展植被调查,记录乔、灌、草的物种、数量、株高、胸径等物种信息,同时按五点式选择5株目标树,分别测量记录距离目标树最近的4株参照树之间的角度与到参照树的距离。根据植被调查的结果,分别计算角尺度、竞争指数和混交度3个林分空间结构指数。角尺度(Wi)可反映林分中林木的水平空间分布格局(惠刚盈等,2016);林分竞争指数(IcI)可以反映目标木在其结构单元中受到的来自其他林木带来的各个方面竞争压力的大小,既能体现出林木所受到的相互遮掩的情况,又可以反映出目标木水平方向上受到的压迫状况(惠刚盈等,2013);混交度(Mi)被定义为目标树i的n株最近邻木中,与目标树不属同种的个体所占的比例,用来描述树种空间隔离程度(惠刚盈等,2016)。计算方式如下:
![]() (1)
(1)
式中:Wi为角尺度;n为最邻近木株数,n=4;Zij表示参照木i与相邻木j所构成的夹角α与标准角α0(α0=72°)的关系,当α>α0时,Zij=0;否则Zij=1。Wi的取值有5种:0、0.25、0.5、0.75、1,分别对应非常均匀、均匀、随机、不均匀、非常不均匀。林分角尺度用林木角尺度均值表示。
 (2)
(2)
式中: IcI为目标树i的竞争指数;Lij为目标树i与竞争木j之间的距离;di为目标树i的胸径;dj为竞争木j的胸径;n为最邻近木株数,n取4。IcI越大,表明树种之间的竞争越激烈;群落竞争指数用单株竞争指数均值表示。
![]() (3)
(3)
式中: Mi为目标树i的混交度;n为邻近木株数;Vij为离散变量。当目标树i与第j株最近邻木非同种时Vij=1, 同种时Vij=0,对林分计算平均混交度。
生物多样性
选取叶厚度( LL,cm)、叶鲜重(LMf,g)、叶饱和鲜重(LMsf,g)、叶干重(LMd,g)和叶面积(LA,cm2)5个植物叶功能性状;茎鲜重(BMf,g)、茎干重(BMd,g)、茎体积(BV,cm3)3个茎性状;根鲜重(RMd,g)、根干重(RMd,g)、根体积(RV,cm-3)、根表面积(RA,cm2)和总根长(RL,cm)5个根性状。利用公式计算比叶面积(SLA(cm2∙g-1)= LA / LMd)、叶干物质含量(LDMC(g∙g-1) = LMd /LMsf)、叶组织密度(LTD(g∙cm-3) = LMd / (LA×LT))、叶含水量(LWC(%)= ((LMf -LMd) / LMf ×100%)、茎干物质含量(BDMC(g∙g-1)=BMd/BMf)、木材密度(WD(g∙cm3)=BMd/BV)、根组织密度(RTD(g·cm-3)=EMd/RV)、比根面积(SRA(cm2∙g-1)=RA/RMd)、比根长(SRL(cm·g-1)=RL/RMd)。每个物种采集9片成熟、健康的叶片,用于叶功能性状的测算。叶片采集完毕后,每个乔木和灌木物种挖掘细根2条,选择相对完整的一条;草本植物整株采集后挑选完整细根一条,用于细根功能性状的测定。利用花枝剪截取树冠外围的2段非当年生小枝,用于小枝功能性状的测算。为保持叶片和枝条的水分饱和,室内功能性状测定均在样本采回8小时内完成。根、茎、叶鲜重采用精度为0.0001的天平称取,叶厚度采用经度为0.01的数显游标卡尺测量,茎体积采用量筒排水法得到。利用CanoScan LiDE 400 扫描仪扫描叶片和细根图像,然后用 Image J软件计算得出叶面积,采用根系图像分析软件Win RHIZO(Pro 2009b)对扫描后的根系进行细根形态特征分析得到根体积、根表面积和总根长。将叶片用清水浸泡24 h称取叶饱和鲜重。然后将所有样品带回实验室,放入75 ℃烘箱烘干至恒重,称取根、茎、叶干重。
根据植被调查数据,选择Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数3种α多样性指数计算群落物种多样性,公式为:
Shannon-Wiener多样性指数: ![]() (4)
(4)
Margalef丰富度指数: ![]() (5)
(5)
Pielou均匀度指数: ![]() (6)
(6)
式中:Pi为种i的个体数占总个体数的比例,S为群落中物种数,N为所有物种的个体总数。
根据功能性状计算群落加权平均值(CWM)两个功能多样性指数。群落加权平均值(CWM)表示群落内某功能性状的平均值,常用来评估群落动态和生态系统特征。
![]() (7)
(7)
式中,n表示样方内树种的物种数;pi表示物种i在样方内的相对多度,traiti表示物种i的性状值。
生态系统服务
选取水源涵养、土壤保肥和产品供给分别表征调节、支持和供给服务。在样地内利用环刀法采集0~10 cm厚度的土壤,分层取土,每5 cm 为一层,现场称取土壤鲜重(SMf)。带回实验室,浸泡12 h后称取饱和土壤持水重(SMs),将土壤放入105℃的烘箱恒温干燥48 h后去除称土壤干重(SMd),利用公式计算土壤容重、毛管持水量、毛管孔隙度等。按五点取样法取样方内0-10 cm表层土,然后均匀混合5个样点的土壤;带回实验室剔除石块和植物根茎叶,过0.149 mm的土样用于测全量养分。每个样地内分别划定1个1 m×1 m的枯落物和草本样方,收获样方内所有枯落物和草本植物。枯落物称取鲜重后置于清水中浸泡24 h称取持水重,草本植物称取鲜重计算草本生物量。
土壤容重采用环刀法测定(林大仪, 2005);土壤饱物理性质采用浸泡法和烘干法测定。现场称取鲜重,带回实验室后浸泡12 h,充分浸水后将环刀置于吸水纸上静置2 h后称吸水重,然后将称完重的环刀置于115 ℃的烘箱中24 h烘干后称干重,最后计算土壤物理性质。土壤化学性质的测定,参考(鲍士旦, 2000),土壤pH值的测定采用水浸提—电位法(国标GB7859-87);有机质含量采用国标GB7857-87的重铬酸钾氧化—外加热法;全氮采用国标GB7173-87 半微量开氏法;土壤全磷的测定采用氢氧化钠碱熔—钼锑抗比色法;土壤全钾的测定:用火焰光度法;实验操作细节和公式还参照鲍士旦等人方法进行计算(鲍士旦, 2000)。
木材产品供给
依据树种、树高、胸径查找贵州省地方标准主要树种单株立木地上二元生物量表(贵州标准化公共服务平台,2022)计算乔木和灌木单株立单株立木材积量,对样地内的单木材积累加得到样地材积量(kg),进而换算为立方米/公顷(t·hm-2)。
土壤保肥:包括土壤碳(C)、氮(N)、磷(P)、钾(K)固持服务
![]() (10)
(10)
![]() (11)
(11)
式中,Sn为第i类养分保持实物量(t),Ai为第i类林分类型面积,为400(m2);Sd为森林土壤平均深度(m),Sd=0.3 m(张俞,2020);Sm为单位体积土壤质量(t/m3);Ni为单位土壤重第i类养分含量(g/kg);1000为单位转换系数;sw为环刀土质量(g);Sv为环刀土体积,为98.125(cm3)。
水源涵养:包括枯落物持水和土壤持水。将所提取的凋落物各组分的样品精确称重(L1)后,置于烘箱中在80℃下烘至恒重后称重(L2)。再将烘干后的凋落物装入尼龙网袋置于水中浸泡24 h,取出枯落物将其空干(以无水滴滴下为标准)后称重(L3),计算凋落物的最大持水量((LCM,g)和最大持水率(LRM,%)。公式如下:
![]()
土壤理化性质与持水性测定及计算方法参见中华人民共和国林业行业标准——森林土壤水分—物理性质的测定(LY/T1215-1999)(国家林业局,1999):
![]() (14)
(14)
式中,SW为土壤最大持水量;Sc为土壤持水重;Sd为土壤干重。
固碳释氧:根据中华人民共和国国家标准——森林生态系统服务功能评估规范(GB/T 38582-2020)(国家林草局,2020)分别核算固碳和释氧。
 (15)
(15)
式中,Gv为植被固碳实物量(t);Rc为CO2中碳含量,为27.27%;Ai为第i类森林类型面积,为400(m2);P为林分净生产力(t·hm-2);Gs为土壤固碳实物量(t);Sd为森林土壤平均深度(m);Sm为单位土壤质量(t/m3);Ci为单位土壤中碳含量(g/kg);Go为植物释氧实物量(t)。
数据处理
利用单因素方差分析(One-way ANOVA)分析不同林型之间林分结构、生物多样性和生态系统服务之间的差异性,并在0.05水平条件下进行显著性检验。利用冗余分析(RDA)分析林分结构和生物多样性对生态系统服务变异的贡献率与相关性,使用MS office excel 2016软件作图。
结果与分析
林分结构差异
研究区乔木、灌木、草本植物共106种77科43属。其中阔叶混交林49种43属23科;阔叶纯林9种9属7科;针阔混交林54种47属32科;针叶林26种24属17科。阔叶混交林主要乔木物种有核桃、桑树、李、云南松、华山松、火棘等;阔叶纯林为核桃树;针阔混交林主要物种有马尾松、白栎、椴树、银白杨等;针叶林中华山松占85%以上,包含少量马尾松、云南松和其他阔叶树种。
从图1 可知,针阔混交林的平均角尺度(0.56)和平均竞争指数(1.75)最大,平均混交度则为阔叶混交林(0.54)最大。说明4种林型都处于随机分布状态,针阔混交林的竞争强度比其他林型高出了48%-304%,针叶林的种间隔离程度最低。角尺度和混交度在不同林型间无显著差异,针阔混交林的竞争指数显著大于其他林型。由于阔叶纯林只有一个乔木物种,混交度为零。
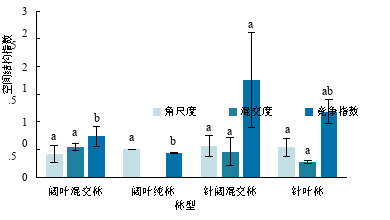
图1 不同林型空间结构指数
生物多样性差异
图2显示,不同林型间物种多样性存在显著差异。其中Shannon多样性指数表现为针阔混交林(3.09)>针叶林(1.93)>阔叶混交林(1.92),Margalef丰富度指数(6.17)和Pielou均匀度指数(0.87)都是针阔混交林最大,其次为阔叶混交林(3.16)和(0.66),针叶林最小(2.97)和(0.80)。针阔混交林的Shannon多样性和Margalef丰富度与其他林型存在显著差异,其他林型间物种多样性差异不显著。阔叶纯林只有一个物种,无法计算物种多样性指数,故为空值。

图2 不同林型物种多样性指数
生态系统服务差异
图4显示,针叶林的木材供给在4种林型中最大(139.10),针阔混交林次之(114.50),阔叶纯林最小(21.25)。针叶林与针阔混交林、阔叶混交林和阔叶纯林之间差异不显著,但是针叶林与针阔混交林都显著高于阔叶混交林和阔叶纯林。

图3 不同林型林木产品供给
如图3所示,4种林型中土壤C、N、P、K固持量均存在显著差异。针阔混交林(9.48)和针叶林(7.62)的C固持量显著大于另外两种林型,阔叶纯林(4.44)最小。N固持量依次为针阔混交林>阔叶混交林>针叶林>阔叶纯林,针阔混交林与其他林型有显著差异,其他林型间差异不显著。P固持量为阔叶纯林的0.16大于阔叶混交林0.14的大于针叶林的0.10大于针阔混交林的0.08,4种林型间均存在显著性差异。针叶林(1.74)的K固持量最大,其次为阔叶纯林(1.04),针阔混交林最小(0.26)。
图4 不同林型土壤养分固持服务
由表2可知,不同林型调节服务差异显著。从水量调节来看,针叶林的土壤最大持水量在4种林型中最大,较其他林型高出了68%~120%。枯落物最大持水量依次为针叶林(8.79±2.55)>针阔混交林(7.07±0.61)>阔叶混交林(17.09±1.09)>阔叶纯林(18.45±4.69)。其中针叶林和针阔混交林显著大于阔叶混交林和阔叶纯林,但是这两种林型间差异不显著。从气候调节来看,针叶林和针阔混交林的固碳释氧量都显著高于阔叶混交林和阔叶纯林,且与水量调节服务一样,针叶林和针阔混交林之间、阔叶纯林和阔叶混交林之间差异不显著。表明在水分拦截和小气候两个方面,针阔混交林和针叶林较另外两种林型拥有更强的调节作用。
表2 不同林型水量与大气调节服务
森林持水 | 林型 | |||
阔叶混交林 | 阔叶纯林 | 针阔混交林 | 针叶林 | |
土壤持水量/t·hm-2 | 313.54±34.04b | 368.71±66.99b | 410.57±29.89ab | 690.05±216.12a |
枯落物最大持水量/t·hm-2 | 8.79±2.55b | 7.07±0.61b | 17.09±1.09a | 18.45±4.69a |
固碳量/t | 5.37±1.23b | 9.62±0.11ab | 11.49±3.99a | 10.07±3.91a |
释氧量/t | 1.66±1.09b | 1.01±0.57b | 5.45±1.62a | 6.627±0.30a |
空间结构对服务的影响
空间结构与服务的冗余分析(图5)显示,第1轴和第2轴的解释变量分别为44.63%和5.24%,总解释变量为49.87%。土壤P、K固持量与林分结构呈显著负相关;混交度和竞争指数与枯落物最大持水量、林木产品供给、固炭释氧显著正相关,与其他服务显著负相关;竞争指数与除土壤P、K固持量外的服务呈显著正相关,表明竞争指数是森林生态系统服务最重要的驱动因子,混交度次之。

图5 空间结构与服务的冗余分析
生物多样性对服务的影响
从生物多样性与生态系统服务的冗余分析(图6)可知,轴1和轴2分别解释了变异的57.64%和22.08%,总解释变量为79.72%。土壤P固持量与叶组织密度显著正相关,与其他服务呈显著负相关。土壤C、N固持量与比叶面积、比根面积、叶干物质含量、多样性、丰富度、均匀度显著正相关,与叶组织密度、茎干物质含量和木材密度显著负相关。土壤K固持量与茎干物质含量、木材密度、根组织密度、多样性、均匀度、丰富度显著正相关,与其他性状加权均值负相关。结果说明物种多样性是供给和调节服务的重要驱动因子,土壤养分固持与生物多样性的关系因养分元素而定。

图 6生物多样性与服务的冗余分析
讨论
本文对比了石漠化治理森林4种林型的空间结构指数、生物多样性指数和生态系统服务差异,并分析了结构、生物多样性与服务之间的相关关系。结果显示,针叶林的木材供给服务在4种林型中最大,其次为针阔混交林,阔叶纯林最小。原因是针叶林中华山松占据了绝对优势,群落平均胸径达到了18.26 cm,较阔叶混交林、阔叶纯林和针阔混交林的胸径分别高出了198%、105%和45%。虽然针阔混交林物种多样性显著高于其他林型,但是群落中乔木占比低于针叶林,且乔木树种林分密度要小于针叶林。而阔叶混交林中大多是灌木,乔木数量较少,因此木材产量较少。而阔叶纯林由人工种植而成,株距通常在5 m以上,且种植时间较短,所以木材产量在4种林型中最少。
土壤C、N、P、K是植物生长发育的重要物质来源,影响植物对养分的吸收、利用等一系列生物生化过程(Qiao et al.,2020)。能够反映土壤碳蓄积动态及养分供应能力,是衡量森林生态系统物质循环和能量流动的重要指标之一(Aerts and Berendse,1989;曾冬萍等,2013)。在相同气候区域的不同森林类型中,地上植被吸收养分元素的差异以及凋落物质量和数量以及分解速率的差异,都将影响土壤养分在土壤中的分布特征(贺若阳等,2016)。针阔混交林的土壤C、N固持量在4种林型中最高,因为针阔混交林的物种多样性最多,凋落物量大,分解速度快,养分归还量大,有利于土壤养分的积累(李静鹏,2014),因此针阔混交林土壤中作为结构性元素的C和光合元素N显著高于其他林型。而阔叶纯林物种数量和种类都较其他林型少, 群落组成相对简单,养分归还量较小,从而使土壤养分“供”大于“还”,导致土壤肥力质量较低(李静鹏,2014)。这也导致枯落物种类和数量低于其他林型,从而限制了土壤C和N的来源。阔叶纯林中土壤P最高,可能是因为受到了人工施肥的影响。阔叶纯林是为石漠化治理而种植的核桃人工林,部分核桃林下还种植了农作物,农户会给地里施加磷肥。此外,由于物种单一,吸收的P元素也较少,所以核桃纯林的土壤P固持量显著高于其他林型。针叶林中土壤K元素积累最多,
森林生态系统一方面通过其繁茂的林冠层实现对大气降水的再分配和有效调节,另一方面森林生态系统内发达的灌草层、致密的林地枯枝落叶层和疏松深厚的土壤层对水分过程产生调蓄作用,整体打造了截获和贮蓄降水的优越环境,发挥着森林植被特有的水源涵养功能(贺淑霞等,2011)。同时,森林生态系统在调节全球碳平衡、减缓大气中CO2等温室气体浓度上升以及维护全球气候等方面中具有不可替代的作用(周玉荣等,2000)。本研究中,针叶林的释氧量和水源涵养服务在4种林型中都显著高于其他林型,针阔混交林的固碳量最高,阔叶纯林的释氧量和枯落物持水量最低,固碳量和土壤持水量则是阔叶混交林最低。因为针阔混交林的土壤C含量和生产力较高,同时,呼吸作用也较强。而针叶林虽然生产力最高,但是叶面积较小,形成的光合产物没有阔叶物种多,但是呼吸作用叶相对弱。因此针阔混交林固碳量显著高于其他林型,但是释氧量较针叶林低。阔叶混交林和阔叶纯林虽然叶面积较针叶林大,但是群落内生物量要少的多,光合产物与枯落物远低于针阔混交林和针叶林,因此固碳释氧量都显著低于针阔混交林和针叶林。森林植被群落的生产力越高,枯枝落叶量越大,土壤微生物数量越多(李勇等,2012)。而针叶林叶纤维发达、木质素含量高,叶片含有油脂等特点使其更难分解(Huang et al.,2007),林下积累了大量未分解的枯落物,所以针叶林的枯落物持水要高于针阔混交林。阔叶纯林枯落物数量少,因此枯落物持水能力最弱。随着林地郁闭度、凋落物产量增加,而林地光热条件的改善和凋落物层的保护作用降低了地表蒸发和地表径流,从而提高了土壤持水能力(李宗勋等,2020)。针叶林的林分密度在4种林型中最大,郁闭度最高,枯落物产量叶最高,同时,叶面积小,蒸腾作用较弱,对土壤水分起到了保护作用。其他林型较高的蒸腾作用和植物需水量吸收了大量土壤水分,降低了土壤持水量。
结论
针阔混交林的空间结构最合理,种间竞争也最为激烈。针阔混交林的生物多样性综合水平最高,其中,Shannon多样性、Margalef丰富度和Pielou均匀度分别为3.09、6.17和0.86。针叶林的产品供给服务、水源涵养服务、释氧服务最好。针阔混交林的固碳服务和土壤养分固持服务供给能力最强。林分结构因子中,竞争指数是服务的最主要驱动因素,生物多样性因子中则是Pielou均匀度指数。针阔混交林拥有更好的生态修复作用,生态脆弱区应该扩大该类森林面积以提高生态系统整体功能。

客服QQ:30444492琼网文【2021】1550-113号
增值电信业务经营许可证:琼B2-20210322
出版物经营许可证:新出发龙华出字第(2021)009号
广播电视节目制作经营许可证:(琼)字第00779号
版权所有 ©2002-2024 期刊网(www.qikanchina.com) 琼ICP备2021005105号



